Известно, что высокочастотная ультрадианная ритмика (табл. 6.1) играет важную роль в функционировании организма. Так, наиболее высокой частотой (60-100 Гц) отличаются нервные импульсы (Беритов, 1959; Коган, Чараян, 1972), затем следуют колебания ЭЭГ с частотой от 0,5 до 70 Гц (Уолтер, 1966; Забродин, Лебедев, 1977; Ливанов, 1984). Декасекундные ритмы были зарегистрированы в биопотенциалах мозга (Богданов и др., 1972; Аладжалова, 1979) и стали основанием для гипотезы о медленной управляющей системе мозга (Аладжалова, 1979). К этому диапазону относятся и колебания биоэлектрохимической активности головного мозга (Швец-Тэнэта-Гурий, 1980), пульса, дыхания, перистальтики кишечника (Баевский, Семенова, Чернышев, 1975; Судаков, 1983). В частности, показано, что биоэлектрическая активность мышц, частота сердечных сокращений и дыхания, амплитуда и частота движений изменяются в среднем через каждые 55 с (Смирнов, Навакатиян, 1980; Леонтьев, 1987). Декаминутные (90 мин) ритмы были открыты в мозговых механизмах ночного сна, которые были названы медленно — и быстроволновой, или парадоксальной, фазами, при этом именно на вторую фазу приходятся сновидения, непроизвольные движения глаз (Ашофф, 1984; Аршавский, 1994). Такой же ритм в последующем был обнаружен в сверхмедленных колебаниях биопотенциалов бодрствующего мозга, связанных с временной динамикой внимания, бдительности оператора (Аладжалова, 1979). Циклы сходной длительности характерны для общей двигательной активности, работоспособности (Kripke, 1972), а также для функций внутренних органов: желудка (Курыгин, 1994), экскреторной функции почек (Lavie, Kripke, 1977).
Однако не изученным остается ультрадианная ритмика развития болевых поведенческих реакций при болевом стрессе у крыс, а также влияние на данный процесс низкоинтенсивного ЭМИ КВЧ, что и явилось задачей данного исследования.
Для решения поставленной задачи экспериментальных животных разделили на три группы (рис. 6.1). Животных первой группы (Кф) подвергали подкожной инъекции в тыльную поверхность стопы задней конечности физиологического раствора. У животных второй (ФТ) и третьей групп (КВЧ+ФТ) тоническую боль вызывали путём подкожной инъекции 5%-ного раствора формалина в дорсальную поверхность стопы задней конечности крыс. Непосредственно перед инъекцией формалина животные третьей группы (КВЧ+ФТ) подвергались превентивному воздействию ЭМИ КВЧ на затылочно-воротниковую область в течение 30-ти минут. Показателями боли служила продолжительность (с) болевой и неболевых (двигательная активность и пассивное поведение) поведенческих реакций за 120 минут наблюдения после инъекции формалина.
Результаты косинор-анализа показали, что у животных всех экспериментальных групп набор и продолжительность периодов в ультрадианном диапазоне не отличались (табл. 6.2).
Однако у животных, подвергнутых изолированному действию болевого стресса (ФТ) во всех выявленных периодах отмечалось достоверное изменение амлитудно-фазовых характеристик изученных показателей относительно значений у животных контрольной группы, подвергнутых инъекции физраствора (табл. 6.3 – 6.5; рис. 6.2). Так, у животных данной группы в ритмике продолжительности болевой реакции отмечалось значительное увеличение амплитуд выделенных периодов относительно значений в контроле (рис. 6.2). Причем с увеличением периодов наблюдалось и увеличение их амплитуд. При этом максимальный рост амплитуд отмечался в ≈7-ми и ≈40-минутных периодах (в 15,73 раза (р<0,01) и в 11,70 раза (р<0,001) соответственно), а минимальный – в периоде ≈5,0 минут (в 1,45 раз (р>0,05)) по сравнению со значением у животных контрольной группы (Кф) (табл. 6.3; рис. 6.2, 6.3).
Величины амплитуд ритмики продолжительности неболевых поведенческих реакций во всех регистрируемых периодах у животных второй группы (ФТ), напротив, достоверно уменьшились. Так, максимальное снижение амплитуды ритмов продолжительности двигательной активности зарегистрировано в периоде ≈7,0 минут (на 92,28 %; р<0,001), а минимальное – в периоде ≈16,0 минут (на 61,79 %, р<0,01) относительно значений у животных первой группы (Кф) (табл. 6.4; рис. 6.2, 6.3). Достоверное уменьшение амплитуд пассивного поведения регистрировалось в периодах ≈7,0; ≈9,0; ≈11,5, ≈15,0 и ≈26,5 минут (в среднем на 48,13±5,18 %; р<0,01) по сравнению со значениями у животных первой группы (Кф).
Таблица 6.2.
Выявленные ультрадианные ритмы показателей продолжительности поведенческих реакций у крыс при экспериментально вызванной тонической боли в «формалиновом тесте»
| Продолжительность выявленных периодов (в минутах) | Показатели | ||
| Болевая реакция | Двигательная активность | Пассивное поведение | |
| ≈2m,5 | – | – | + |
| ≈3m,0 | + | + | – |
| ≈5m,0 | + | + | + |
| ≈7m,0 | – | + | – |
| ≈7m,5 | + | – | – |
| ≈9m,0 | – | + | + |
| ≈11m,0 | + | – | – |
| ≈11m,5 | – | – | + |
| ≈12m,5 | – | + | – |
| ≈14m,5 | + | – | – |
| ≈15m,0 | – | – | + |
| ≈16m,0 | – | + | – |
| ≈21m,5 | + | – | – |
| ≈22m,0 | – | + | – |
| ≈26m,5 | – | – | + |
| ≈36m,0 | – | + | – |
| ≈40m,0 | + | – | – |
| ≈63m,0 | – | + | – |
Примечание: «+» – наличие периода данной продолжительности, «–» – отсутствие.
Таблица 6.3.
Данные косинор-анализа продолжительности болевой реакции у крыс, подвергнутых введению физиологического раствора (Кф), изолированному (ФТ) и комбинированному с воздействием низкоинтенсивного электромагнитного излучения крайне высокой частоты (КВЧ+ФТ) действию болевого фактора в «формалиновом тесте» (![]() )
)
| Показатели | Экспериментальные группы воздействия | ||
| Кф(1) | ФТ (2) | КВЧ+ФТ (3) | |
| Период (минуты) | 3,03±0,47 | 3,02±0,31 | 2,87±0,39 |
| Амплитуда (усл. ед.) | 0,16±0,10 | 2,56±0,34
Р1,2<0,01 |
1,33±0,32
Р1,3<0,05 Р2,3<0,05 |
| Фазы (радианы) | 12,60±1,25 | 2,27±0,40
Р1,2<0,001 |
4,39±0,67
Р1,3<0,001 Р2,3<0,05 |
| Период (минуты) | 5,60±0,10 | 4,90±0,41 | 4,920±0,34 |
| Амплитуда (усл. ед.) | 2,22±0,15 | 3,22±0,67 | 3,016±0,53 |
| Фазы (радианы) | 1,61±0,77 | 3,67±0,53
Р1,2<0,05 |
1,004±0,48
Р2,3<0,01 |
| Период (минуты) | 7,50±0,36 | 7,78±0,32 | 6,85±0,65 |
| Амплитуда (усл. ед.) | 0,85±0,40 | 4,86±0,34
Р1,2<0,001 |
3,97±0,10
Р1,3<0,001 Р2,3<0,05 |
| Фазы (радианы) | 1,37±0,08 | 3,88±0,23
Р1,2<0,001 |
2,02±0,13
Р1,3<0,01 Р2,3<0,001 |
| Период (минуты) | 11,20±0,51 | 10,05±0,05 | 11,00±0,30 |
| Амплитуда (усл. ед.) | 1,67±0,03 | 3,31±0,21
Р1,2<0,001 |
1,61±0,32
Р2,3<0,01 |
| Фазы (радианы) | 2,20±0,29 | 2,43±0,21 | 1,19±0,40 |
| Период (минуты) | 14,07±0,83 | 14,30±0,42 | 14,60±0,78 |
| Амплитуда (усл. ед.) | 1,67±0,31 | 4,69±0,20
Р1,2<0,001 |
3,12±0,44
Р1,3<0,05 Р2,3<0,01 |
| Фазы (радианы) | 0,66±0,48 | 2,44±0,07
Р1,2<0,01 |
0,42±0,31
Р2,3<0,001 |
| Период (минуты) | 21,05±1,326 | 21,72±1,17 | 21,63±0,07 |
| Амплитуда (усл. ед.) | 1,09±0,01 | 5,65±0,46
Р1,2<0,001 |
1,22±0,14
Р2,3<0,001 |
| Фазы (радианы) | 0,91±0,24 | 0,63±0,29 | 0,40±0,01 |
| Период (минуты) | 34,55±3,17 | 42,22±4,98 | 39,67±2,67 |
| Амплитуда (усл. ед.) | 0,62±0,32 | 7,19±0,49
Р1,2<0,001 Р2,3<0,001 |
3,15±0,52
Р1,3<0,01 Р2,3<0,001 |
| Фазы (радианы) | 0,80±0,10 | 0,54±0,16 | 0,37±0,10
Р1,3<0,05 |
Примечание: р1-3 достоверность различий по критерию Стьюдента при сравнении с данными групп, обозначенными в таблице 1-3 соответственно.
Таблица 6.4.
Данные косинор-анализа продолжительности двигательной активности у крыс, подвергнутых введению физиологического раствора (Кф), изолированному (ФТ) и комбинированному с воздействием низкоинтенсивного электромагнитного излучения крайне высокой частоты (КВЧ+ФТ) действию болевого фактора в «формалиновом тесте» (![]() )
)
| Показатели | Экспериментальные группы воздействия | ||
| Кф(1) | ФТ (2) | КВЧ+ФТ (3) | |
| Период (минуты) | 3,07±0,27 | 3,03±0,20 | 2,83±0,30 |
| Амплитуда (усл. ед.) | 2,93±0,73 | 0,86±0,10
Р1,2<0,05 |
2,24±0,41
Р2,3<0,01 |
| Фазы (радианы) | 4,42±0,56 | 4,32±1,13 | 4,26±0,74 |
| Период (минуты) | 5,00±0,22 | 5,02±0,32 | 5,30±0,41 |
| Амплитуда (усл. ед.) | 4,89±0,22 | 0,92±0,20
Р1,2<0,001 |
3,36±0,53
Р1,3<0,01 Р2,3<0,001 |
| Фазы (радианы) | 1,63±0,34 | 3,90±0,46 | 1,96±0,28
Р1,3<0,01 |
| Период (минуты) | 7,15±0,43 | 6,95±0,35 | 7,20±0,21 |
| Амплитуда (усл. ед.) | 5,20±0,69 | 0,40±0,08
Р1,2<0,001 |
2,19±0,19
Р1,3<0,05 Р2,3<0,01 |
| Фазы (радианы) | 2,56±0,48 | 3,71±0,18
Р1,2<0,05 |
1,58±0,45
Р2,3<0,01 |
| Период (минуты) | 8,67±0,57 | 8,96±0,28 | 9,50±0,30 |
| Амплитуда (усл. ед.) | 6,72±0,98 | 0,89±0,16
Р1,2<0,001 |
2,20±0,16
Р1,3<0,01 Р2,3<0,001 |
| Фазы (радианы) | 2,56±0,48 | 3,43±0,31
Р1,2<0,05 |
3,07±0,09
Р1,3<0,05 Р2,3<0,001 |
| Период (минуты) | 13,15±0,35 | 12,12±0,49 | 12,27±0,37 |
| Амплитуда (усл. ед.) | 6,49±1,30 | 1,64±0,27
Р1,2<0,01 |
6,72±0,41
Р2,3<0,001 |
| Фазы (радианы) | 2,50±0,25 | 1,50±0,15
Р1,2<0,01 |
2,30±0,11
Р2,3<0,01 |
| Период (минуты) | 16,07±0,79 | 15,65±0,56 | 16,50±0,52 |
| Амплитуда (усл. ед.) | 3,46±0,40 | 1,32±0,26
Р1,2<0,01 |
3,85±0,40
Р2,3<0,001 |
| Фазы (радианы) | 1,54±0,37 | 0,93±0,28 | 1,78±0,02
Р2,3<0,05 |
| Период (минуты) | 22,01±1,34 | 22,42±1,12 | 21,88±1,09 |
| Амплитуда (усл. ед.) | 11,38±2,41 | 1,46±0,21
Р1,2<0,001 |
4,01±0,37
Р1,3<0,01 Р2,3<0,001 |
| Фазы (радианы) | 1,21±0,22 | 1,21±0,16 | 1,54±0,11 |
| Период (минуты) | 36,90±3,42 | 36,15±2,63 | 35,83±2,94 |
| Амплитуда (усл. ед.) | 10,16±0,96 | 1,67±0,23
Р1,2<0,001 |
6,56±0,60
Р1,3<0,01 Р2,3<0,001 |
| Фазы (радианы) | 0,80±0,07 | 0,89±0,09 | 0,92±0,06 |
| Период (минуты) | 62,30±0,69 | 63,20±5,92 | 64,07±0,47 |
| Амплитуда (усл. ед.) | 11,41±2,00 | 1,97±0,40
Р1,2<0,001 |
5,00±0,29
Р1,2<0,01 Р2,3<0,001 |
| Фазы (радианы) | 0,54±0,00 | 0,56±0,05 | 0,53±0,02 |
Примечание: р1-3 достоверность различий по критерию Стьюдента при сравнении с данными групп, обозначенными в таблице 1-3 соответственно.
Таблица 6.5.
Данные косинор-анализа продолжительности пассивного поведения у крыс, подвергнутых введению физиологического раствора (Кф), изолированному (ФТ) и комбинированному с воздействием низкоинтенсивного электромагнитного излучения крайне высокой частоты (КВЧ+ФТ) действию болевого фактора в «формалиновом тесте» (![]() )
)
| Показатели | Экспериментальные группы воздействия | ||
| Кф(1) | ФТ (2) | КВЧ+ФТ (3) | |
| Период (минуты) | 2,88±0,21 | 2,61±0,30 | 2,52±0,25 |
| Амплитуда (усл. ед.) | 4,34±0,45 | 3,18±0,42 | 7,26±0,60
Р1,3<0,01 Р2,3<0,001 |
| Фазы (радианы) | 5,44±0,40 | 2,90±0,57
Р1,2<0,05 |
5,82±0,83
Р2,3<0,05 |
| Период (минуты) | 4,58±0,30 | 5,08±0,34 | 4,74±0,26 |
| Амплитуда (усл. ед.) | 5,33±0,47 | 4,85±1,01 | 5,24±1,28 |
| Фазы (радианы) | 1,47±0,57 | 1,36±0,70 | 1,37±0,46 |
| Период (минуты) | 6,75±0,28 | 6,97±0,29 | 6,70±0,40 |
| Амплитуда (усл. ед.) | 9,45±0,44 | 3,24±0,58
Р1,2<0,001 |
7,82±0,85
Р2,3<0,01 |
| Фазы (радианы) | 2,09±0,93 | 1,15±0,11 | 2,54±0,22
Р2,3<0,001 |
| Период (минуты) | 8,78±0,31 | 8,63±0,20 | 9,20±0,31 |
| Амплитуда (усл. ед.) | 11,15±0,65 | 5,29±0,92
Р1,2<0,01 |
6,39±0,42
Р1,3<0,01 |
| Фазы (радианы) | 0,86±0,25 | 1,40± 0,22 | 1,54±0,19 |
| Период (минуты) | 11,13±0,55 | 10,93±0,38 | 12,20±0,32 |
| Амплитуда (усл. ед.) | 9,04±0,64 | 5,42±1,09
Р1,2<0,01 Р2,3<0,001 |
16,19±0,47
Р1,3<0,001 Р2,3<0,001 |
| Фазы (радианы) | 0,89±0,04 | 1,19±0,19 | 0,94±0,03
Р1,3<0,001 Р2,3<0,05 |
| Период (минуты) | 14,60±0,80 | 15,00±0,74 | 15,73±0,59 |
| Амплитуда (усл. ед.) | 9,94±1,52 | 6,34±0,66
Р1,2<0,05 |
11,42±1,64
Р2,3<0,01 |
| Фазы (радианы) | 1,09±0,07 | 1,08±0,13 | 0,85±0,11
Р1,3<0,02 |
| Период (минуты) | 25,27±2,11 | 27,86±3,16 | 26,28±2,79 |
| Амплитуда (усл. ед.) | 11,97±0,67 | 6,45±1,70
Р1,2<0,05 Р2,3<0,05 |
11,63±1,00
Р2,3<0,05 |
| Фазы (радианы) | 0,50±0,18 | 0,83±0,21 | 0,48±0,16 |
Примечание: р1-3 достоверность различий по критерию Стьюдента при сравнении с данными групп, обозначенных 1-3 в таблице соответственно.
|
|
|
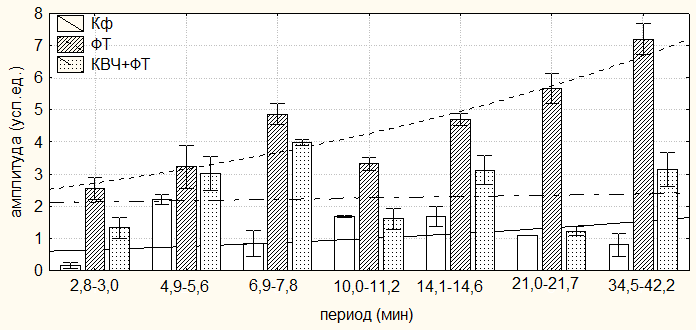
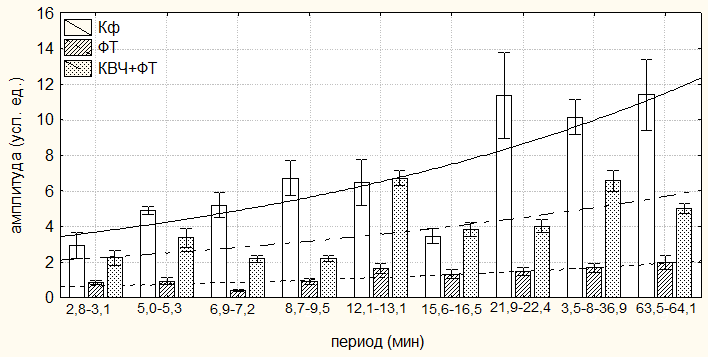
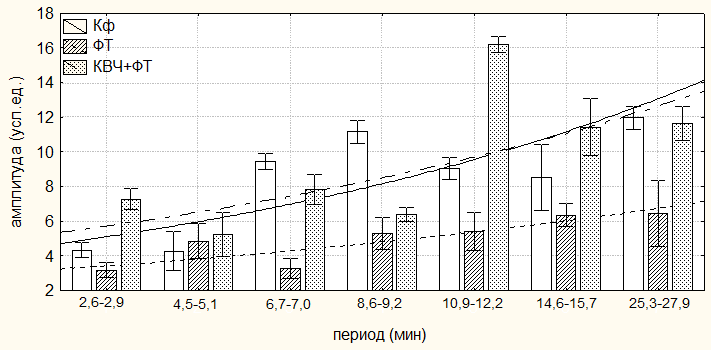
Рис. 6.2. Амплитуды периодов ультрадианного ритма продолжительности болевой (А) и неболевых (двигательной активности (Б) и пассивного поведения (В)) поведенческих реакций у крыс, подвергнутых введению физиологического раствора (0,9 % раствор NaCl) (Кф), изолированному (ФТ) и комбинированному с воздействием низкоинтенсивного электромагнитного излучения крайне высокой частоты (КВЧ+ФТ) действию болевого фактора в «формалиновом тесте» при экспериментально вызванной тонической боли.
А
|
|
||
|
|||
|
![]()
![]()
![]()
|
|
|
|
|
|
Б
|
|||
|
|||
|
|||
|
|||
|
|
|
|
|
|
В
|
|
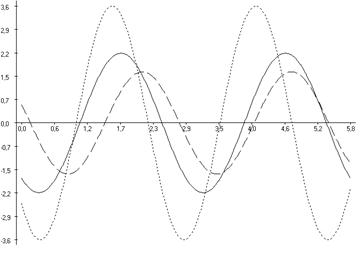
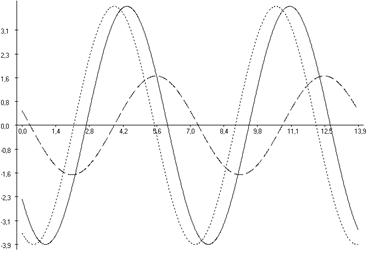
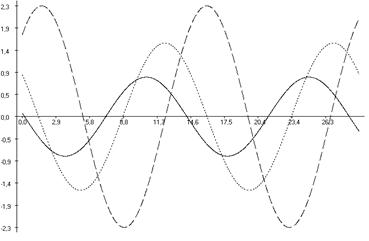
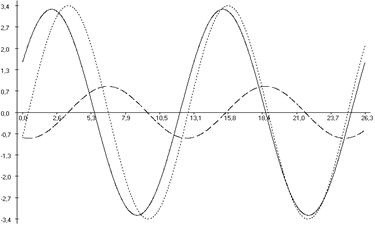
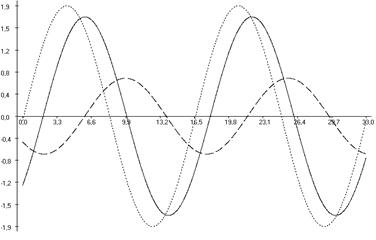
Рис. 6.3. Фазовые соотношения ультрадианной ритмики продолжительности болевой реакции (А), двигательной активности (Б) и пассивного поведения (В) у крыс, подвергнутых введению физиологического раствора (1), изолированному (2) и комбинированному с однократным воздействием низкоинтенсивного электромагнитного излучения крайне высокой частоты (3) действию болевого фактора в «формалиновом тесте».
Кроме того во всех выделенных периодах у крыс второй группы, подвергнутых изолированной инъекции формалина (ФТ) отмечались фазовые смещения исследуемых показателей относительно таковых у животных контрольной группы (Кф). Так, наибольший достоверный фазовый сдвиг длительности болевой реакции был отмечен в периоде продолжительностью ≈3,0 минут (на 232,04о; р<0,001), наименьший – в периоде продолжительностью ≈5,0 минут (на 117,85о; р<0,05) относительно значений соответствующих показателей у крыс в первой группе. При этом в периодах ≈11,0; ≈21,5 и ≈40,0 минут наблюдалась близость фаз (p>0,05) относительно значений у крыс контрольной группы (табл. 6.3; рис. 6.2, 6.3).
В ультрадианной ритмике продолжительности двигательной активности максимальное смещение фаз отмечалось в ≈5,0-минутном периоде (на 104,84о; р<0,05). При этом в периодах продолжительностью ≈3,0; ≈16,0; ≈22,0; ≈36,0 и ≈63,0 минут достоверных фазовых изменений не наблюдалось (табл. 6.4). В ритмике показателя длительности пассивного поведения достоверное смещение фаз отмечено лишь в периоде ≈2,5 минуты (на 145,53о; р<0,01) относительно данных у крыс первой группы (табл. 6.5, рис. 6.3).
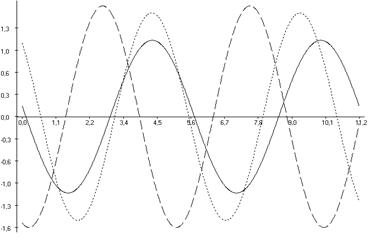
Таким образом, полученные результаты свидетельствуют о том, что при действии болевого стресса у животных наблюдалось существенное нарушение ультрадианной ритмики продолжительности как болевых, так и неболевых (двигательной активности и пассивного поведения) поведенческих реакций, выраженные в значительном изменении амплитуд (увеличение или уменьшение) и сдвигах фаз в выделенных периодах относительно значений у животных контрольной группы, что свидетельствует о развитии десинхроноза. Кроме того, косинорограммы интегральных ультрадианных ритмов показателей продолжительности болевой реакции, двигательной активности и пассивного поведения свидетельствуют об ослаблении координации между поведенческими реакциями у животных, подвергнутых экспериментальной тонической боли, что указывает на развитие внутреннего десинхроноза, который является следствием развития в организме стресс-реакции (рис. 6.4).
У животных третьей группы, которые подвергались действию болевого фактора на фоне КВЧ-воздействия отмечалось достоверное изменение амплитудно-фазовых характеристик относительно значений соответствующих показателей у животных второй группы (ФТ), подвергнутых изолированной инъекции формалина. Так, имело место уменьшение амплитуд продолжительности болевой реакции во всех выделенных периодах (табл. 6.3; рис. 6.2). Максимальное снижение данного показателя было отмечено в периоде ≈21,5 минут (на 78,41 %; р<0,001), а минимальное – в периоде ≈5,0 минут (на 6,38 %; р>0,05).
При этом отмечался существенное увеличение амплитуд как продолжительности двигательной активности (максимальное – в периоде продолжительностью ≈7,0 минут (на 444,46 %; р<0,01), минимальное – в периоде ≈63,5 минут (на 153,98 %; р<0,001)), так и пассивного поведения (максимальное – в ≈11,5-минутном периоде (на 198,70 %; р<0,001), минимальное – в периоде ≈5,0 минут (на 7,94 %; p>0,05)) во всех выделенных периодах относительно значений данного показателя у животных второй группы (ФТ) (табл. 6.3, 6.4, рис. 6.2, 6.3).
|
|
|
|
|
|
|
|
|
|
|
|
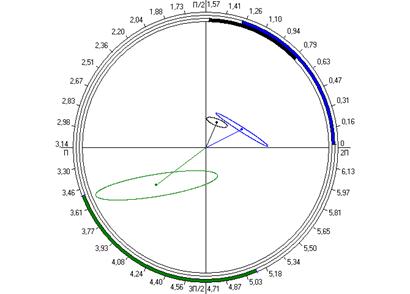
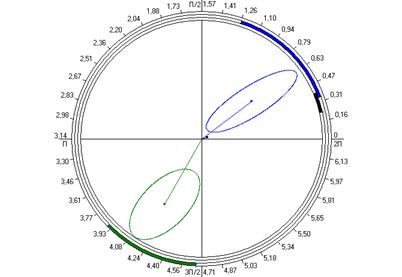
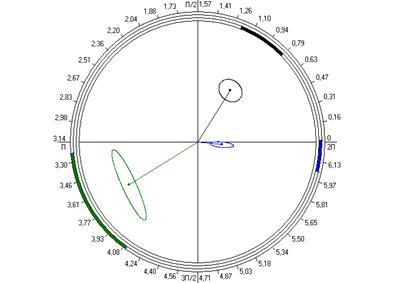
Рис. 6.4. Косинорограммы интегральных ульрадианных ритмов показателей продолжительности болевой (1) и неболевых (двигательная активность (2) и пассивное поведение (3)) поведенческих реакций у крыс, подвергнутых введению физиологического раствора (0,9 % раствор NaCl) (Кф), изолированному (ФТ) и комбинированному с однократным воздействием низкоинтенсивного электромагнитного излучения крайне высокой частоты (КВЧ+ФТ) действию болевого фактора в «формалиновом тесте» при экспериментально вызванной тонической боли в периоде ≈5,0 минут.
При этом амплитуды изученных поведенческих реакций во всех выделенных периодах в большой степени достоверности приближались к таковым у крыс контрольной группы (табл. 6.3 – 6.5; рис. 6.2)
В ультрадианной ритмике продолжительности поведенческих реакций у животных третьей группы также отмечались определенные фазовые изменения в выделенных периодах относительно значений данных показателей у крыс второй группы (ФТ). Так, в ритмике продолжительности болевой реакции максимальное смещение фаз регистрировалось в ≈5,0-минутном периоде (на 152,55о; р<0,01), минимальное – в периоде ≈7,5 минут (на 107,09о; р<0,001) относительно значений у животных второй группы (ФТ), подвергнутых действию только болевого фактора в «формалиновом тесте» (табл. 6.3; рис. 6.2, 6.3). Близость фаз отмечалась в периодах ≈11,0; ≈21,5 и ≈40,0 минут (табл. 6.3).
Фазовые смещения зарегистрированы также в длительности неболевых поведенческих реакций. Наиболее выраженный фазовый сдвиг продолжительности двигательной активности отмечен в ≈9,0-минутном периоде (на 122,188о; р<0,01). При этом в периодах ≈3,0; ≈5,0; ≈22,0; ≈36,0 и ≈63,0 минут наблюдалась близость фаз. Максимальный фазовый сдвиг продолжительности пассивного поведения зарегистрирован в периоде ≈2,5 минут (на 167,54о; р<0,05), тогда как в периодах ≈5,0; ≈9,0; ≈15,0 и ≈26,5 минут отмечена близость фаз относительно значений этих показателей у крыс второй группы.
При этом значения фаз выделенных периодов продолжительности болевой и неболевых поведенческих реакций у крыс, предварительно подвергавшихся КВЧ-воздействию с большой степенью достоверности приближались к таковым (в основном на уровне тенденции) у крыс, которым вместо формалина вводили физраствор (табл. 6.3 – 6.4).
Таким образом, при дополнительном воздействии низкоинтенсивного ЭМИ КВЧ выявлена способность данного физического фактора модифицировать ультрадианную ритмику длительности болевой и неболевых поведенческих реакции у крыс при действии болевого стресса, что проявляется в большем сходстве амплитудно-фазовых характеристик данных показателей с таковыми у животных контрольной группы, подвергнутых ложному действию болевого фактора – инъекции физраствора, чем у животных, также подвергнутых инъекции формалина, но без предварительного КВЧ-воздействия. Косинорограммы интегральных ультрадианных ритмов показателей поведенческих реакций у животных, подвергнутых комбинированному действию ЭМИ КВЧ и болевого стресса наглядно иллюстрируют восстановление и нормализацию фазовых взаимоотношений данных показателей, что свидетельствует о внутренней синхронизации продолжительности всех поведенческих реакций (рис. 6.4). По-видимому, при нарушении ультрадианной ритмики при болевом стрессе воздействие низкоинтенсивного ЭМИ КВЧ оказывает коррегирующее, синхронизирующее действие, что еще раз доказывает антистрессорное действие миллиметрового излучения.
Автор — Джелдубаева Эльвиза Рашидовна