Согласно нашим (Чуян, 1992; 2004) и литературным (Алисов и др., 1989; Пясецкий и др., 1991; Ельский, 1997; Темурьянц и др., 2001; Лушников и др., 2001) данным, существует зависимость биологической эффективности низкоинтенсивного ЭМИ КВЧ от индивидуально-типологических особенностей организма человека и животных. Желание иметь доступный и удобный критерий для оптимизации индивидуальных параметров воздействия ЭМИ КВЧ привело к многочисленным экспериментальным и клиническим исследованиям, в которых использовались различные подходы. Одним из объективных методов, позволяющих выделить индивидуальные различия между животными и сформировать однородные экспериментальные группы является тест ОП (Hаll, 1934). В настоящее время этот тест применяется в различных экспериментальных исследованиях и позволяет относительно быстро выявить индивидуальные различия между животными, оценить целостную физиологическую реакцию животного на новую обстановку, включающую элементы двигательного, ориентировочно-исследовательского и эмоционального поведения (Маркель, 1981; Кулагин и др., 1986; Семагин и др., 1988; Буреш и др., 1991; Гуляева и др., 1997). Показано, что у животных с различными индивидуально-типологическими особенностями поведения в тесте ОП имеются существенные различия в характере энергетического, белкового и других видов метаболизма (Гернштейн, 1991; Егорова и др., 1995; Онуфриев, 1995; Гуляева, Степаничев, 1997), активности окислительно-восстановительных ферментов (Краковский, 1987), перекисного окисления липидов (Левшина, Гуляева, 1995; Степаничев, 1995), моноаминергических систем (Jeste, 1980; Arons, 1992; Герцова, 1993; Кругликов и др., 1995) и других физиологических и биохимических показателей.
Ведущую роль в процессах адаптации организма к различным условиям среды, в том числе ЭМИ, играет головной мозг. Однако мозг не является равновесной системой, его активность обеспечивается функциональной асимметрией, которая выступает как общая фундаментальная закономерность деятельности ЦНС человека и животных, а выраженность асимметрии определяет адаптивность организма (Симмерницкая, 1978; Бианки, 1989). В настоящее время накоплено значительное число фактов, свидетельствующих о наличии латеральной специализации мозга человека и животных (Bradshaw, 1990). Отмечено, что асимметрия может проявляться на анатомическом, сенсорном, когнитивном или моторном уровнях, а ее характер зависит от гормонального статуса, уровня развития и средовых влияний. Адекватными методами для исследования моторной асимметрии у животных, которая отражает асимметрию ЦНС, т. е. доминирование правого или левого полушария головного мозга является тест ОП и Т-образного лабиринта (Бианки, 1980).
В наших предыдущих исследованиях было показано. что как уровень двигательной активности в тесте ОП, так и профиль моторной асимметрии животных могут служить критерием чувствительности к низкоинтенсивному ЭМИ КВЧ (Чуян, 1992; Чуян и др., 2004). Однако зависимость аналгетического действия ЭМИ КВЧ от индивидуальных особенностей животных не изучена. В связи с этим задачей данной серии экспериментов явилось выявление зависимости выраженности аналгетического эффекта низкоинтенсивного ЭМИ КВЧ от индивидуальных особенностей животных.
Для реализации поставленной задачи была проведена серия экспериментов (табл. 5.1; рис. 5.6) с крысами, имеющими разный уровень двигательной активности: НДА, СДА и ВДА и низкую эмоциональность, которые определялись в тесте ОП.
Анализ величины коэффициента асимметрии в тестах ОП и Т-образного лабиринта и уровня двигательной активности в тесте ОП показал, что среди крыс со СДА преобладают «правши» (Кас=51,15+1,97 усл. ед.), среди животных c ВДА – «левши» (Кас=35,83+0,53 усл. ед.), а крысы с НДА преимущественно являются «амбидекстрами» (Кас=0,50+0,05 усл. ед.).
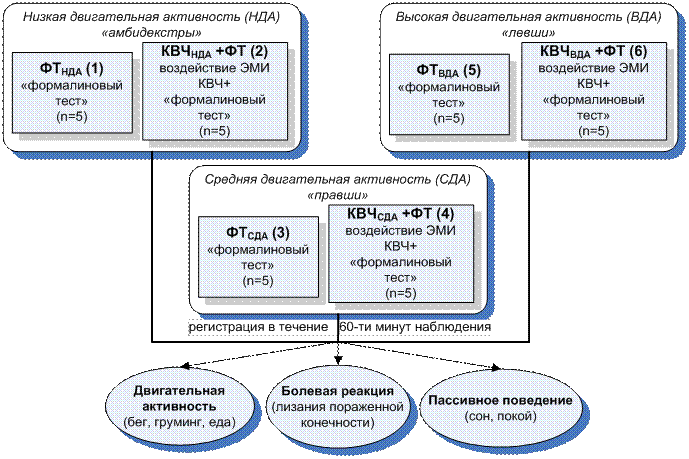
Рис. 5.6. Схема организации экспериментальных исследований по изучению зависимости антиноцицептивного эффекта низкоинтенсивного электромагнитного излучения крайне высокой частоты (КВЧ) от индивидуальных особенностей животных с низким (НДА), средним (СДА) и высоким (ВДА) уровнем двигательной активности при экспериментально вызванной тонической боли в «формалиновом тесте» (ФТ).
Предвартельно отобранных животных разделили на шесть групп по пять особей в каждой (рис. 6.6) и подвергали воздействию болевого стресса в «формалиновом тесте». Животных 2-ой (КВЧНДА+ФТ), 4-ой (КВЧСДА+ФТ) и 6-ой (КВЧВДА+ФТ) групп непосредственно перед введением формалина предварительно подвергали КВЧ-воздействию (λ = 7,1 мм, ППМ = 0,1мВт/см2, экспозиция – 30 минут, локализация – затылочно-воротниковая область). Интенсивность протекания болевой реакции оценивали по продолжительности болевой и неболевых (двигательная активность и пассивное поведение) поведенческих реакций в течение 60-ти минут наблюдения после инъекции формалина.
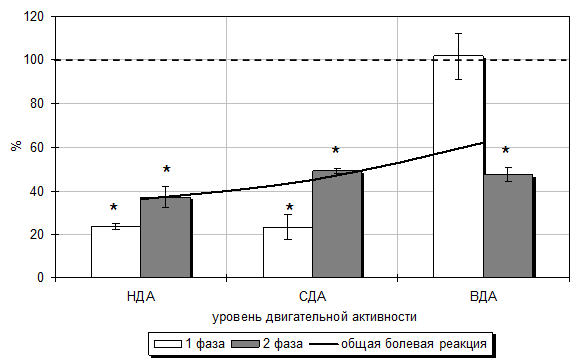
Как показали результаты проведенного исследования, продолжительность поведенческих реакций у крыс с разным уровнем двигательной активности и профилем моторной асимметрии в «формалиновом тесте» была различна. Так, у крыс третьей группы (ФТСДА) со СДА и правосторонней моторной асимметрией, которые преобладают в популяции (следовательно, у них развивается наиболее типичная реакция на различные воздействия) введение формалина вызвало бурную болевую реакцию лизания конечности общей продолжительностью 1001,00±20,85 с (первая острая фаза – 158,60±17,10 с; вторая тоническая – 842,40±17,57 с). Продолжительности неболевых поведенческих реакций представлены в табл. 5.6.
У крыс с НДА, у которых моторная асимметрия не выражена («амбидекстры») продолжительности болевой и неболевых поведенческих реакций были недостоверно ниже таковых у крыс со СДА (табл. 5.6).
У животных с ВДА и преобладающей левосторонней моторной асимметрией продолжительность болевой реакции имела тенденцию к увеличению (на 3,43 %; р>0,05) относительно значений у животных третьей группы (ФТСДА). При этом длительность двигательной активности была больше на 143,70 % (р<0,001), а пассивного поведения – на 11,61 % (р<0,001) относительно значений у животных со СДА, также подвергнутых инъекции формалина (табл. 5.6).
Таблица 5.6.
Общая продолжительность (с) болевой и неболевых поведенческих реакций у крыс с низким (НДА), средним (СДА) и высоким (ВДА) уровнями двигательной активности и разным профилем моторной асимметрии при изолированном (ФТ) и комбинированном с воздействием низкоинтенсивного электромагнитного излучения крайне высокой частоты (КВЧ+ФТ) действием болевого фактора в «формалиновом тесте» (![]() )
)
| Типы поведенче-ских реакций | Экспериментальные группы животных | |||||
| НДА («амбидекстры») | СДА («правши) | ВДА («левши») | ||||
| ФТ(1) | КВЧ+ФТ(2) | ФТ(3) | КВЧ+ФТ(4) | ФТ(5) | КВЧ+ФТ(6) | |
| Болевая реакцияВ том числе: | 947,75±44,20P1,2<0,001P1,4<0,001
P1,6<0,001 |
341,33±13,67P1,2<0,001P2,3<0,001
P2,4<0,001 P2,5<0,001 P2,6<0,001 |
1001,00±20,85P2,3<0,001P3,6<0,001 | 449,00±11,79P1,4<0,001P2,4<0,001
P3,4<0,001 P4,5<0,001 P4,6<0,001 |
1035,33±36,67P2,5<0,001P4,5<0,001
P5,6<0,001 |
643,50±19,53P1,6<0,001 p2,6<0,001P3,6<0,001
P4,6<0,001 P5,6<0,001 |
| 1 фаза | 187,00±10,33 p1,2<0,01P1,4<0,01 | 44,33±2,88P1,2<0,01P2,3<0,01
P2,5<0,001 P2,6<0,001 |
158,60±17,10P2,3<0,01P3,4<0,01 | 37,00±8,14P1,4<0,01P3,4<0,01
P4,5<0,001 P4,6<0,001 |
175,00±16,65 p2,5<0,001 | 178,00±6,12 p4,5<0,001 p4,6<0,001 p2,6<0,001 |
| 2 фаза | 799,92±104,35 p1,2<0,001P1,4<0,001P1,6<0,001 | 297,00±3,61P1,2<0,001P2,3<0,001
P2,4<0,001 P2,5<0,001 P2,6<0,01 |
842,40±17,57 p3,4<0,01 p3,6<0,001 | 412,00±9,02P1,4<0,001P2,3<0,001
P2,4<0,001 P3,4<0,01 P4,5<0,001 |
860,33±23,40 p2,5<0,001P4,5<0,001 p5,6<0,001 | 406,25±26,82P1,6<0,001P2,6<0,01
P3,6<0,001 p5,6<0,001 |
| Бег | 173,67±19,43 p1,2<0,001P1,4<0,001P1,5<0,001 | 337,00±12,49P1,2<0,001P2,3<0,01
P2,4<0,001 P2,6<0,001 |
191,80±22,43P2,3<0,01P3,4<0,01
P3,5<0,01 |
605,33±25,08P1,4<0,001P2,4<0,001
P3,4<0,01 P4,5<0,001 p4,6<0,001 |
330,33±5,84P1,5<0,001P3,5<0,01 p4,5<0,001 p5,6<0,001 | 132,67±18,68P2,6<0,001P4,6<0,001 p5,6<0,001 |
| Груминг | 68,00±15,87 p1,2<0,05P1,4<0,001P1,6<0,01 | 233,67±55,84P1,2<0,05P2,3<0,01 | 62,20±5,36P2,3<0,01P3,4<0,001
P3,6<0,01 |
196,00±17,02P1,4<0,001P3,4<0,001 | 109,33±40,81 | 210,00±38,55P3,6<0,01P1,6<0,01 |
| Прием пищи | 0,00±0,00P1,3<0,001P1,5<0,001 | 23,00±11,79 p2,3<0,05P2,5<0,001 | 52,80±5,28P1,3<0,001P2,3<0,05
P3,5<0,001 P3,6<0,001 |
35,00±23,71P4,5<0,001 | 308,00±43,32P1,5<0,001P2,5<0,001
P3,5<0,001 P4,5<0,001 p5,6<0,001 |
0,00±0,00P3,6<0,001 p5,6<0,001 |
| Сон | 818,33±429,24 | 834,00±194,87 p2,3<0,01P2,4<0,01 | 241,60±11,16P2,3<0,01P3,4<0,001
P3,5<0,001 P3,6<0,05 |
0,00±0,00P3,4<0,001 p4,5<0,001P4,6<0,01
P2,4<0,01 |
1083,33±142,82 p3,5<0,001P4,5<0,001 p5,6<0,05 | 531,33±156,47P3,6<0,05 p4,6<0,01P5,6<0,05 |
| Покой | 3240,00±199,79P1,3<0,01 p1,4<0,01P1,5<0,05
P1,6<0,05 |
3572,00±109,07P2,3<0,01 p2,4<0,01P2,5<0,001 | 3850,60±21,37P1,3<0,01P2,3<0,01
P3,5<0,001 |
4114,67±25,69P1,4<0,01P2,4<0,01
P3,4<0,001 |
2533,67±146,95P1,5<0,05P2,5<0,001
P3,5<0,001 p4,5<0,001 p5,6<0,001 |
3848,00±149,35P1,6<0,05P4,5<0,001
P5,6<0,001 |
Примечание: р1-6 – достоверность различий по критерию Стьюдента при сравнении с данными групп, обозначенными в таблице 1 – 6.
Таким образом, не выявлено различий в продолжительности болевой реакции в «формалиновом тесте» у животных разных типологических групп. Однако у животных с ВДА в тесте ОП отмечались более высокие значения продолжительности двигательной активности и в «формалиновом тесте», что свидетельствует об адекватности применения этих тестов для оценки двигательного и эмоционального поведения.
Под влиянием ЭМИ КВЧ интенсивность болевой реакции значительно снижалась, что согласуется с результатами наших исследований представленных в главе 4. Однако степень снижения продолжительности этих реакций у крыс в «формалиновом тесте» на фоне воздействия ЭМИ КВЧ зависела от уровня двигательной активности и моторной асимметрии, что подтверждает данные ряда исследований о том, что один и тот же фактор у различных животных может вызвать различные ответные реакции (Юматов, Мещерякова, 1990; Сантана Вега, 1991; Чуян, 1992; Грабовская, 1992; Судаков, 1998). Так, у животных четвертой группы (КВЧСДА+ФТ) со СДА и правосторонней моторной асимметрией после однократного 30-тиминутного воздействия ЭМИ КВЧ наблюдалось уменьшение продолжительности болевой реакции на 55,14 % (р<0,001) (длительности первой острой фазы – на 76,67 % (р<0,001), второй тонической – на 51,09 % (р<0,001)) относительно значений данных показателей у животных со СДА, подвергнутых изолированной инъекции формалина. При этом продолжительность неболевых поведенческих реакций увеличилась: двигательной активности –на 172,60 % (р<0,001), а пассивного поведения –– на 0,55 % (р>0,05) по сравнению с животными, которые подвергались только действию болевого фактора (табл. 5.6; рис. 5.7).
У крыс второй группы (КВЧНДА+ФТ) с НДА ЭМИ КВЧ вызвало даже более выраженное, чем у животных со СДА уменьшение продолжительности болевой реакции – на 63,98 % (р<0,001) (первой острой фазы – на 76,29 % (р<0,001), а второй тонической – на 62,87 % (р<0,001)) относительно значений соответствующих показателей у животных с НДА, подвергнутых только инъекции формалина. При этом общая продолжительность болевой реакции была меньше на 34,54 % (р<0,001) по сравнению со значениями у животных со СДА, также подвергнутых КВЧ-воздействию, что проявилось в основном за счет уменьшения продолжительности второй фазы болевой реакции (на 52,18 %; р<0,001).
|
|
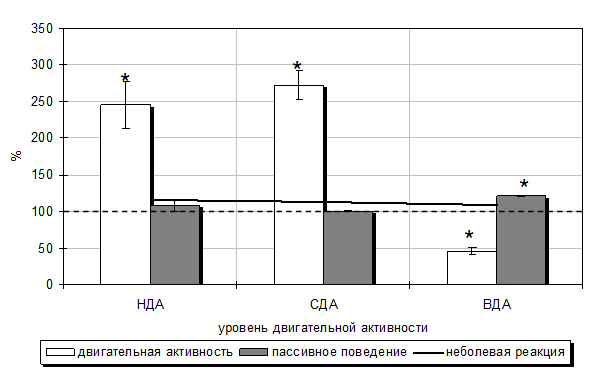
Рис. 5.7. Изменение продолжительности болевой (первой и второй фаз) (А) и неболевых (двигательная активность и пассивное поведение) (Б) поведенческих реакций в «формалиновом тесте» у крыс с низкой (НДА), средней (СДА) и высокой (ВДА) двигательной активностью, подвергнутых комбинированному воздействию низкоинтенсивного электромагнитного излучения крайне высокой частоты и болевого фактора (в % относительно значений у животных тех же фенотипических групп, подвергнутых изолированному действию болевого фактора).
* – достоверность различий относительно значений показателей у животных, подвергнутых изолированному воздействию болевого стресса.
При этом увеличились длительности как двигательной активности (на 145,66 %; р<0,001), так и пассивного поведения (на 8,57 %; р>0,05) по сравнению с животными с НДА, которые подвергались только действию болевого фактора (табл. 5.6; рис. 5.7). Относительно значений данных показателей у животных четвертой группы (КВЧСДА+ФТ) продолжительность двигательной активности уменьшилась на 29,02 % (р<0,01), а пассивного поведения, напротив, увеличилась на 7,08 % (р<0,01).
Таким образом, под влиянием предвартельного КВЧ-воздействия у животных с НДА и СДА («амбидекстры» и «правши»), подвергавшихся болевому воздействию отмечалось увеличение двигательной активности и уменьшение пассивного поведения в «формалиновом тесте».
Увеличение двигательной активности у животных данных фенотипических групп при воздействии ЭМИ КВЧ свидетельствует об уменьшении реакции страха, затаивания и позволяет говорить о возрастании возбудимости ЦНС, что свидетельствует о повышении неспецифической резистентности животных к болевому стрессу, поскольку известно, что активная резистентность реализуется благодаря высокой функциональной активности защитных систем и преобладания в мозгу процессов возбуждения в физиологических пределах (Симонов, 1981).
Иная реакция на КВЧ-воздействие развивалась у животных «левшей» с ВДА. У крыс этой группы КВЧ-воздействие также привело к уменьшению общей продолжительности болевой реакции на 37,85 % (р<0,001) относительно значений этого показателя у крыс с ВДА, которые не подвергались облучению ЭМИ мм диапазона. Однако снижение болевой чувствительности произошло только за счет уменьшения длительности второго тонического компонента болевой реакции (на 52,78 %; р<0,01). При этом продолжительность первой острой фазы болевой реакции у крыс с ВДА в отличие от животных других фенотипических групп не изменилась и соответствовала таковой у крыс с ВДА, которые дополнительно не подвергались КВЧ-воздействию (рис. 5.7 – А).
Таким образом, у крыс с ВДА и левосторонней моторной асимметрией зарегистрировано увеличение продолжительности болевой реакции на 43,32 % (р<0,001) относительно значений у крыс со СДА и на 88,53 % (р<0,001) по сравнению с животными с НДА.
Иная направленность отмечалась и в динамике продолжительности неболевых поведенческих реакций у животных с ВДА после КВЧ-воздействия: продолжительность двигательной активности уменьшилась на 54,17 % (р<0,01), а пассивного поведения – повысилась на 21,08 % (р<0,001) относительно значений у животных пятой группы (ФТВДА) (табл. 5.6; рис. 5.7).
Уменьшение двигательной активности у животных этой фенотипической группы под влиянием болевого фактора свидетельствует о развитии торможения в ЦНС и эмоциональной реакции страха. Следовательно, ЭМИ КВЧ у животных с ВДА и левосторонней асимметрией препятствовало развитию болевого стресса, однако обладало меньшей аналгетической эффективностью по сравнению с его применением у крыс с другими индивидуально-типологическими особенностями.
Полученные результаты показали, что изменения продолжительности поведенческих реакций у животных под влиянием сочетанного действия ЭМИ КВЧ и болевого стресса зависят от индивидуально-типологических особенностей животных. При этом максимальный аналгетический эффект ЭМИ КВЧ отмечен у животных с НДА («амбидестров»), минимальный – с ВДА («левшей»). Данные этого исследования согласуются с результатами наших предыдущих исследований, в которых показано, что наиболее выраженное антистрессорное действие при гипокинетическом стрессе под влиянием миллиметровых волн отмечается у крыс «амбидекстров» с НДА (Чуян, 2004), которые характеризуются пассивно-оборонительной формой поведения, что свидетельствует о слабом тормозном типе нервной системы (Хоничева, 1984). Наименее выраженное антистрессорное действие зарегистрировано у животных-«левшей» с ВДА, характеризующихся активно-оборонительным характером поведения, что позволяет отнести их к сильному неуравновешенному типу нервной системы (Маркель, 1981; Симонов, 1984). Следовательно, различие в аналгетических эффектах КВЧ-воздействия, отмеченное в данном исследовании у крыс может быть обусловлено силой нервных процессов в ЦНС.
Таким образом, уровень двигательной активности и профиль моторной асимметрии могут служить критерием индивидуальной чувствительности к низкоинтенсивному ЭМИ КВЧ.
В исследованиях, представленных в этой главе выявлена выраженная зависимость антиноцицептивного действия низкоинтенсивного мм излучения от физических параметров ЭМИ (наличия или отсутствия поляризации), продолжительности однократного и курсового воздействия, а также индивидуально-типологических особенностей организма животных. При этом изменение характеристик ЭМИ КВЧ не вызывало качественно новых изменений поведенческих реакций у крыс в болевых тестах, а лишь усиливало или ослабляло антиноцицептивный эффект этого физического фактора.
Автор — Джелдубаева Эльвиза Рашидовна